
O kwasach nukleinowych:

 |
| Johann Friedrich Miescher |
1. Występowanie kwasów nukleinowych.
Każda komórka, każdego organizmu na Ziemi zawiera DNA i RNA. Co więcej komórki - oprócz "własnego" materiału genetycznego mogą zawierać również pasożyty wewnątrzkomórkowe, które także posiadają własny materiał genetyczny. Są to np. wirusy, które podzielić możemy na wirusy DNA i wirusy RNA (tzw. retrowirusy). Do komórki przenikać mogą także zakaźne cząsteczki RNA, które nazywamy wiroidami.
Przedstawmy miejsca występowania DNA i RNA za pomocą tabeli.
MIEJSCA WYSTĘPOWANIA:

2. Funkcje kwasów nukleinowych.
Zadaniem kwasów nukleinowych jest przechowywanie informacji genetycznej i pośredniczenie w produkcji białek zgodnie z zasadami kodu genetycznego. Kwasy nukleinowe mogą pełnić także funkcje enzymów (ta rola przez bardzo długi czas przypisywana była tylko i wyłącznie białkom). Dziś doskonale zdajemy sobie sprawę z tego, że rybozymy, spełniają funkcje analogiczne do enzymów białkowych, a nawet - dzięki tej funkcji, RNA jest w stanie podlegać autokatalizie. Rybozymy występują u wszystkich organizmów i biorą udział w syntezie białek i przemianach kwasów nukleinowych. Przykładem jest np. transferaza peptydylowa wchodząca w skład rybosomów, która katalizuje tworzenie wiązań peptydowych pomiędzy aminokwasami w trakcie procesu translacji. Rybozymy wycinają również introny ze świeżo zsyntezowanego pre-mRNA.
| Źródło: https://slideplayer.pl/slide/417857/ - "Aktywność katalityczna enzymów". |
W latach 1979-1982 Thomas Cech prowadził badania procesu wycinania intronów, czyli tzw. splicingu na jednokomórkowym eukarioncie - Tetrahymena thermophila. Badania trwały bardzo długo - Cech bowiem nie mógł znaleźć za żadne skarby białka odpowiedzialnego za wycinanie fragmentów niekodujących. Bo i takiego białka nie było! To właśnie wtedy nastąpiło dojście do wniosku, że RNA podlega autokatalizie - sam wycina zbędne fragmenty. Jest to rzecz nieprawdopodobna - tak zaskakująca dla biologów, że niektórym ciężko było w nią uwierzyć. W 1989 r. Thomas Cech i Sidney Altman otrzymali za swą pracę nagrodę Nobla w dziedzinie chemii.

Thomas Cech jest chemikiem amerykańskim. W dzieciństwie ojciec - fizyk, zainteresował go nauką. Interesował się w młodości, zwłaszcza geologią. Ponoć w czasach gimnazjum zapukał do drzwi profesorów geologii na University of Iowa i poprosił ich o omówienie struktur krystalicznych, meteorytów i skamieniałości. Interesował się również literaturą, m.in. studiował "Odyseję" Homera i "Piekło" Dantego. Studiował w Berkeley, a dziś od roku 2000 kieruje Instytutem Medycznym Howarda Hughesa. W obszarze jego zainteresowań naukowych znajdował się proces transkrypcji. W latach 70-tych ubiegłego wieku zajmował się badaniem splicingu. Drugą dziedziną badań było badanie struktury i funkcji telomerów i telomerazy. Nagrodę Nobla otrzymał w 1989.

Sidney Altman - to kanadyjsko-amerykański biolog molekularny o polskim pochodzeniu, urodzony 7 maja 1939 roku i tu zwracam uwagę na to, że rodzina Altmanów w odpowiedniej porze opuściła Europę. Jego matka pochodziła bowiem z Białegostoku. Wraz z siostrą przybyła do Kanady w wieku 18 lat i pracowała w fabryce włókienniczej, by zarobić pieniądze na przyjazd reszty rodziny. Jego ojciec, urodzony na Ukrainie, był pracownikiem kołchozu w Związku Radzieckim. Sidney Altman mawiał o swoich rodzicach, iż: "To od nich dowiedziałem się, że ciężka praca w stabilnym otoczeniu może przynieść nagrody, nawet jeśli tylko w nieskończenie małych przyrostach" - istotnie po przyjeździe ojca Altmana do Kanady - rodzice naukowca otworzyli małe przedsiębiorstwo - sklep spożywczy w Montrealu, co pozwoliło im na skromne i szczęśliwe życie z dala od wojennych, a później powojennych zawirowań politycznych. Sidney Altman ukończył studia fizyczne w MIT. Niedługo później zaczął wykładać na Uniwersytecie Yale. Prowadził badania nad rybonukleazami. Splicing, tak jak i w przypadku Cecha mieścił się w kręgu jego zainteresowań. Badania prowadził niezależnie od niego.
Podczas pobytu w Yale, praca Altmana, która przyczyniła się do przyznania mu Nagrody Nobla zawierała analizę katalitycznych właściwości rybozymowej RNazy P , rybonukleoproteinowej cząstki składającej się zarówno z strukturalnej cząsteczki RNA, jak i jednej (u prokariotów ) lub więcej (u eukariotów ) białek. Początkowo uważano, że w kompleksie bakteryjnej RNazy P podjednostka białkowa była odpowiedzialna za aktywność katalityczną kompleksu, który bierze udział w dojrzewaniu tRNA. Podczas eksperymentów, w których kompleks był odtwarzany w probówkach, Altman i jego grupa odkryli, że składnik RNA, w izolacji, był wystarczający do obserwowanej aktywności katalitycznej enzymu, wskazując, że samo RNA miało właściwości katalityczne, co było odkryciem, które przyniosło mu Nagrodę Nobla. Chociaż kompleks RNazy P występuje również w organizmach eukariotycznych, jego późniejsza praca ujawniła, że w tych organizmach podjednostki białkowe kompleksu są niezbędne do aktywności katalitycznej, w przeciwieństwie do bakteryjnej RNazy P.
Obiekt badań Cecha:

Oto i Tetrahymena thermopila - protist zwierzęcy, orzęsek. To ciekawe, że występuje u niego 7 płci, a każda może współżyć z inną. Daje nam to 21 orientacji seksualnych. Jednocześnie nie występuje u nich dymorfizm płciowy (czy więc owe możliwości współżycia aby na pewno słusznie jest określać orientacjami seksualnymi?). Co więcej - płeć determinowana jest u nich przez prawdopodobieństwo, jednakże nie nie tak jak u nas - gdzie heterogametyczność męska determinuje 50% prawdopodobieństwa na urodzenie dziewczynki i 50% prawdopodobieństwa na urodzenie chłopca. U Tetrahymena themopila mamy tzw. gen Mat, a prawdopodobieństwo określamy nie samym przekazaniem allelu warunkującego płeć, a jego działaniem.
3. ROLA RNA:
Cząsteczki RNA pełnią ważne role w funkcjonowaniu komórki. Odpowiadają bowiem za regulację ekspresji genów (miRNA), a także wchodzą w skład aparatu translacyjnego (rRNA tworzy rybosom, tRNA - dobudowuje kolejne aminokwasy do syntezowanego łańcucha polipeptydowego), odszyfrowuje zakodowaną informację genetyczną za pomocą antykodonu. Oprócz tych typowych, znanych RNA, istnieją także:
piRNA - niekodujące cząsteczki RNA, które biorą udział w posttranskrypcyjnych, epigenetycznych mechanizmach wyciszania retranspozonów i innych elementów genetycznych, które związane są z przemieszczaniem się genów w procesie transpozycji. Cząsteczki te wykazują ekspresję w komórkach zwierząt, głównie w męskich komórkach płciowych, w procesie spermatogenezy.
4. Budowa kwasów nukleinowych:
Kwasy nukleinowe są biopolimerami, zbudowanymi z nukleotydów. Monomer składa się z nukleozydu- cząsteczki pentozy (dla RNA - rybozy, a dla dna - deoksyrybozy), do której przy pierwszym atomie węgla, wiązaniem N-glikozydowym przyłączona jest zasada azotowa (puryna lub pirymidyna). Do trzeciego oraz piątego atomu węgla przyłączona jest również reszta kwasu fosforowego (V). Stanowi ona swego rodzaju łącznik dla dwóch sąsiednich pentoz polimeru - uściślając - jest to wiązanie fosfodiestrowe.
Adenina - Puryny - ich cząsteczki wyglądają jak sześciokąt mający wspólny bok z pięciokątem.
Guanina
Tymina - Pirymidyny - ich cząsteczki mają kształt sześciokąta.
Cytozyna

Puryny - powyższe krótkie uogólnienia mogą nasuwać nam na myśl, że skoro struktura puryny to sześciokąt o wspólnym boku z pięciokątem, a pirymidyna to sześciokąt, to puryna zawierać może w sobie pirymidynę. I tak jest w istocie! Puryna to związek składający się z pirymidyny (sześciokąt) i imidazolu (pięciokątna cząsteczka). Imidazol jest dosyć powszechny w cząsteczkach organicznych, np. w wielu enzymach wchodzi w skład centrów aktywnych. Kwas moczowy to produkt metabolizmu zasad purynowych. Pokarm, który spożywamy zawiera kwasy nukleinowe - muszą więc one zostać poddane procesom metabolicznym i w postaci kwasu moczowego, ulec wydaleniu z organizmu. Wówczas gdy występuje podwyższona ilość kwasu moczowego we krwi, mamy do czynienia z dną moczanową. Wówczas konieczne jest ograniczenie ilości puryn w diecie (rezygnuje się głównie z wysokobiałkowych produktów).

Pirymidyny - pirymidyna ma kształt sześciokąta. Zawiera 4 atomy węgla (4 wierzchołki) i 2 atomy azotu (2 wierzchołki).
Podsumowując: obie zasady mają ze sobą wiele wspólnego. Są to związki organiczne zawierające azot o bardzo częstym występowaniu w świecie przyrody. Jednakże co najważniejsze - tworzą one szczebelki cząsteczki DNA, przypominającej drabinę. Sam fakt iż nici DNA są antyrównoległe jest konsekwencją tego w jaki sposób zasady azotowe łączą się ze sobą wiązaniami wodorowymi.
Doskonale jest to widoczne na poniższym obrazku. Zauważ, że w zależności, czy połączymy wiązaniami wodorowymi adeninę z tyminą w kolejności T-A, czy A-T, cząsteczka tyminy i adeniny musi ulec odpowiedniemu obróceniu w celu zainicjowania tworzenia się wiązania wodorowego. Wynika stąd antyrównoległość nici.
 |
| Źródło: https://slideplayer.pl/slide/61050/ |
Możesz także na podstawie powyższych obrazków zauważyć, że zasady łączą się ze sobą w określony sposób. Mianowicie puryna łączy się z pirymidyną. Jednakże nie zawsze łączą się one ze sobą dwoma wiązaniami wodorowymi. Guanina i Cytozyna jest w stanie połączyć się trzema takimi wiązaniami! Jeżeli lubisz kwestionować pewne sprawy, to zapytasz pewnie: "dlaczego?". W końcu u Adeniny odstaje wodór (nie jest to narysowane, bo w pierścieniach i łańcuchach węglowych dla uproszczenia nie dorysowuje się wodorów, wszak każdy wie, że one tam są), a u Tyminy odstaje tlen. Aż prosi się o to, żeby stworzyło się wiązanie pomiędzy nim a wodorem. Jednakże tak się nie dzieje. Różnica odległości jest niewielka od dopuszczalnej, jednakże nie pozwala na to, by wiązanie wodorowe powstało. W przeciwnym razie, faktycznie pomiędzy adeniną i tyminą mogłyby być trzy wiązania. Z kolei w Guaninie i Cytozynie takiego problemu nie ma, bo we wszystkich odstających częściach atomy są na tyle blisko siebie, że wiązania powstają.

Wiązania wodorowe, które występują pomiędzy zasadami, zwane również "mostkami wodorowymi", to słabe wiązania chemiczne, które polegają na oddziaływaniu elektrostatycznym pomiędzy atomem wodoru i atomem elektroujemnym, który zawiera wolne pary elektronowe. Ładunek wodoru oznaczyć możemy jako q+ (dodatni), a ładunek atomu elektroujemnego jako q- (ujemny). Ładunki te przyciągają się wzajemnie. Jest to oczywiście duże uogólnienie, jednakże w ten sposób należy to sobie tłumaczyć. Wiązania wodorowe obecne są np. w ciekłej wodzie. Zauważ, że cząsteczka H20 ma region naładowany ujemnie (0) i dwa regiony o ładunku dodatnim (H2). Zatem pomiędzy cząsteczkami wody występują wiązania wodorowe wpływające na wiele jej właściwości.

A oto fragment polimeru:

Dla B-DNA: kąt skręcenia między sąsiednimi parami zasad: +34,6stopnia ; skok helisy: 3,54nm ; kierunek skręcenia: prawoskrętna; średnica: 2,37 nm. Liczba par zasad przypadająca na skręt helisy: 10,4.
Po opisanym wcześniej odkryciu Fryderyka Mieschera przez prawie 100 lat struktura DNA pozostawała zagadką. Dopiero James Watson i Francis Crick na podstawie zdjęć rentgenowskich Rosalind Franklin (przekazał im je Maurice Wilkins, współpracujący z Rosalind), skonstruowali oni model podwójnej helisy. Zrobili to w 1954r., za co otrzymali Nagrodę Nobla w 1962r. wraz z Mauricem Wilkinsem.
Kwas deoksyrybonukleinowy
U Eukariontów kwas deoksyrybonukleinowy jest zlokalizowany przede wszystkim w jądrze komórkowym (a oprócz tego a autonomicznych organellach takich jak mitochondria i chloroplasty). U Prokariontów znajduje się bezpośrednio w cytoplazmie i występuje w postaci tzw. chromosomu bakteryjnego lub genoforu w obszarze zwanym nukleoidem. Z kolei u Wirusów obecny jest opakowany w kapsyd.
Monomerami DNA są deoksyrybonukleotydy:
dAMP - deoksyadenozynomonofosforan
dGMP - deoksyguanozynomonofosforan
dTMP - deoksytymidynomonofosforan
dCMP - deoksycytydynomonofosforan
Nukleotydy to ufosforylowane nukleozydy. Powyżej mamy do czynienia z monofosforanami, czyli powyższe nukleotydy zawierają jedną grupę fosforanową. Nukleozydem nazywamy zaś zasadę polączoną z cukrem oznaczaną jak dX, np. dA (deoksyadenozyna). Literką P oznaczamy fosforan nieorganiczny. A zatem zapiszmy reakcje fosforylacji deoksynukleozydów:
dA + P ----> dAMP
dG + P ----> dGMP
dC + P ----> dCMP
dT + P ----> dTMP
NUKLEOZYDY:

BUDOWA NUKLEOTYDU (DNA):

DNA jest zwykle dwuniciowy (dsDNA = double strand DNA), zaś jego łańcuchy są (dla przypomnienia) antyrównoległe. Koniec 5'PO4 leży naprzeciwko końca 3'OH drugiej nici. Łańcuchy zwijają się wokół własnej osi i tworzą tym samym prawoskrętną helisę.
Długość DNA i upakowanie:
Po rozpakowaniu DNA z chromosomów i połączeniu cząsteczek w jedną nić, ludzkie DNA w komórce somatycznej ma długość ok. 2 metrów. Trzeba więc je w odpowiedni sposób "upakować". Służą do tego białka histonowe u Eucaryota i białka histonopodobne u Procaryota. U Eucaryota jest możliwe bardzo ścisłe upakowanie w tzw. chromosom metafazowy - najbardziej skondensowaną formę, jaką może przybrać materiał genetyczny. Jednakże w niedzielącej się komórce eukariotycznej DNA występuje w postaci chromatyny, czyli tzw. włókna 30 nm.
Łańcuch DNA zawiera informację o:
- kolejności aminokwasów w białkach,
- o sekwencji licznych RNA niekodujących,
- o regulacji ekspresji genów,
- o sekwencjach o niejasnym znaczeniu (większość jądrowego DNA)
Sekwencja białek kodowana jest w postaci trójek nukleotydowych, odpowiadających odpowiednim aminokwasom i kodonom terminacyjnym, podczas biosyntezy białka.
Informacja genetyczna jest zapisana także poprzez ułożenie nici w nukleosomach (mówi się o tzw. drugim kodzie, zaraz po kodzie genetycznym, czyli kodzie histonowym).
REPLIKACJA DNA (faza S - Synthesis Interfazy w cyklu komórkowym)
Proces skopiowania nic podwójnej DNA (samoodtwarzanie się DNA) to replikacja. Jest procesem półzachowawczym, czyli semikonserwatywnym. W każdej z uzyskanych nici podwójnych DNA jedna nić będzie macierzysta, a jedna nowa. Proces ten zachodzi w trakcie Interfazy w fazie S i jest procesem endoenergetycznym (wymaga dostarczenia energii).
Substraty:
- matryca DNA,
- trifosforany deoksyrybonukleotydów (dNTP) - jako budulec dla nowej nici DNA,
- ATP/CTP/GTP - jako źródło energii dla helikaz
Enzymy biorące udział w replikacji:
1) Topoizomeraza - wprowadza lub usuwa skręty z podwójnej nici DNA przez przerywanie i ponowne łączenie jednej lub obu nici. Topoizomerazy konwertują energię chemiczną pochodzącą od ATP w energię napięcia cząsteczki o strukturze superhelikalnej. Rozplatając DNA udostępniają matrycę dla enzymów replikujących.
2) Helikazy - rozplątują i rozrywają wiązania wodorowe pomiędzy zasadami azotowymi i tym samym powodują rozdzielenie się nici.
3) Primaza - jest odmianą Polimerazy RNA zależnej od DNA. Primaza aktywowana jest przez helikazę DNA i po aktywacji syntezuje na obu niciach DNA krótkie (11+/-1 zasad) komplementarne odcinki startowego RNA, wykorzystywane przez polimerazę DNA do rozpoczęcia syntezy nowych nici DNA w procesie replikacji DNA. Do syntezy RNA nie potrzebuje w przeciwieństwie do polimeraz DNA odcinków starterowych z wolnym końcem 3'-OH. Przy połączeniu z helikazą tworzy kompleks - primosom, czyli tzw. aparat replikacyjny.
4) Polimerazy DNA - katalizują one reakcję łączenia się kolejnych nukleotydów w łańcuch polinukleotydowy komplementarny do pojedynczej nici DNA.
5) Egzonukleaza - usuwa startery RNA z nici.
6) Ligaza DNA - uzupełnia wiązania fosfodiestrowe w szkielecie nowo zsyntezowanej nici DNA.
+ enzymy pomocnicze.
DZIAŁANIE TOPOIZOMERAZY:

Następuje nukleofilowy atak grupy hydroksylowej reszty tyrozyny w centrum katalitycznym topoizomerazy na wiązanie fosfodiestrowe 3'--5' w DNA. Utworzone w ten sposób pęknięcie przeciąga nietkniętą nić. Uwolniona zostaje w ten sposób cząsteczka kwasu deoksyrybonukleinowego i usunięty skręt helisy.
DZIAŁANIE POLIMERAZY DNA:
Polimeraza DNA katalizuje syntezę DNA w czasie replikacji. Synteza polega na polimeryzacji deoksyrybonukleotydów poprzez wytwarzanie wiązań fosfodiestrowych pomiędzy nimi. Substratami są nukleotydy jednofosforanowe. Polimeraza DNA potrzebuje wzorca, dlatego syntezowane są primery (startery) przez primazę - rodzaj polimerazy RNA. Polimeraza DNA wydłuża starter. Robi to przez dobudowywanie nukleotydów komplementarnych do matrycy na końcu 3' startera. Polimeraza posiada aktywność korektorską i może odłączać źle wstawiony nukleotyd.
DZIAŁANIE LIGAZY:
Ligaza wytwarza wiązanie pomiędzy końcem hydroksylowym 3', a końcem fosforowym 5' w reakcji zależnej od adenozynotrifosforanu (ATP). Reakcja katalizowana przez ligazę, nazywana jest ligacją. Ułatwiają ją tzw. lepkie końce, czyli kilkunukleotydowe odcinki DDNA znajdujące się na końcu dwuniciowego DNA, powstające w wyniku niesymetrycznego przecięcia cząsteczki DNA.
np. 5'AGTCTGCGT 5'CGGGCGGCATGCATTG3'
3'TCAGACGCAGCCCGCCG5' 3'TACGTAAC5'
↓ligacja
5'AGTCTGCGTCGGGCGGCATGCATTG3'
3'TCAGACGCAGCCCGCCGTACGTAAC5'
Tworzą się więc w ligacji:
- wiązania wodorowe,
- wiązania fosfodiestrowe
Funkcje ligazy:
- łączenie fragmentów Okazaki (Ligaza DNA I),
- naprawianie uszkodzeń DNA (przerywania nici) (Ligaza DNA III, Ligaza DNA IV)
U Eucaryota ligazy zależne są od ATP.
MECHANIZM REPLIKACJI:
Zasady replikacji są podobne u wszystkich organizmów, przy czym największe różnice występują pomiędzy bakteriami, a archeonami i eukariontami.
U bakterii:
- replikacja zaczyna się w ustalonym miejscu,
- postępuje bardzo szybko (1000 nukleotydów na sekundę),
- zachodzi w jednym miejscu,
U Eucaryota:
- replikacja wolniejsza (ok. 50 nukleotydów na sekundę)
- zachodzi równocześnie w wielu miejscach
Należy pamiętać, że Polimeraza DNA posiada zdolność działania w kierunku od końca 3' do końca 5' (tam dołącza trifosforany deoksyrybonukleotydów), dlatego nić syntezowana jest w kierunku od 5' do 3'.
Z tego powodu nić DNA 3'--->5' jest nicią opóźnioną. Syntezowana jest w przeciwną stronę "Fragmentami Okazaki".
Zobrazowanie replikacji reakcją chemiczną:

UWAGA: Przy strzałce/nad strzałką dopisać powinienem jeszcze: Polimeraza DNA i Mg2+, ponieważ są to elementy niezbędne do zajścia procesu.

Różnice w replikacji u Eucaryota i Procaryota są doskonale widoczne na etapie Inicjacji:

ELONGACJA:
Jedna nić wydłużana jest w sposób ciągły (wiodąca), a druga fragmentami (Fragmenty Okazaki) - nić opóźniona. Każdy fragment Okazaki zaczyna się od syntezy startera, do którego dobudowywane są nukleotydy. Prymaza katalizuje przyłączanie starterów. Następnie wycinane są startery, a nowe fragmenty DNA łączone w ciągłą nić, tzw. nić opóźnioną. Między nukleotydami sąsiadującymi tworzą się wiązania fosfodiestrowe w wyniku działania Ligazy DNA oraz przyłączenia grupy 3'-OH Deoksyrybozy do wiązania między wewnętrzną grupą fosforanową i dwoma zewnętrznymi grupami fosforanowymi sąsiedniego nukleotydu. Następstwem tego jest uwolnienie zewnętrznych grup fosforanowych w postaci nieorganicznego pirofosforanu, który jest produktem ubocznym procesu.
TERMINACJA:
Terminacja replikacji następuje w momencie ukończenia procesów przebiegających jednocześnie w różnych miejscach replikujących się cząsteczek DNA. Do terminacji dochodzi, gdy widełki replikacyjne replikonu natkną się na specjalną sekwencję terminacyjną na końcu liniowego DNA (w telomerze chromosomu).
Po terminacji zachodzą przemiany poreplikacyjne (np. metylacja zasad azotowych). Metylacja odpowiada za wiele procesów związanych z rozwojem organizmów, np. za różnicowanie się tkanek w odpowiedni sposób. Oznacza to, że wpływa na regulację ekspresji genu. Metylacja może np. zahamować ekspresję danego genu, czyniąc go nieaktywnym. Wówczas luźna - aktywna genetycznie struktura euchromatyny, zamianiana jest w mocniej upakowaną heterochromatynę, dużo mniej aktywną genetycznie, lub w ogóle nieaktywną, niedostępną dla czynników transkrypcyjnych. Przykładem takiej poreplikacyjnej metylacji jest proces lionizacji, czyli inaktywacja chromosomu X w celu zamiany go w nieaktywne genetycznie Ciałko Barra. Co więcej metylacja regulująca ekspresję genów udowadnia, że nasz fenotyp nie jest całkowicie uzależniony od genotypu. Zmienność fluktuacyjna w istocie związana jest z metylacją. Także nasze zachowania zdrowotne wpływają na ekspresję naszych genów, ponieważ przyczyniają się do metylacji zasad azotowych. Wspominałem już o tzw. kodzie histonowym - to właśnie przyłączanie grup metylowych reguluje ułożenie nici DNA na oktamerze histonowym - nić przylega do nich bardziej wówczas, gdy przyłączonych jest do niej więcej grup -CH3, a gdy mniej, jest ułożona luźniej.
Problem wolnych zakończeń:
Proces replikacji cząsteczek DNA niekolistych (eukariotycznych) jest powiązany z problemem wolnych zakończeń. Po degradacji primerów nici opóźnionej, primery są degradowane i zastępowane nicią DNA. Przy każdej replikacji miejsce przyczepu najbliższego telomerowi nie może zostać skopiowane, ponieważ polimeraza nie może się cofać. W efekcie replkikacji towarzyszy skracanie chromosomu. Jest to prawdopodobnie przyczyna starzenia się. Skracaniu zapobiega obecność telomerazy, która przeprowadza odwrotną transkrypcję tych odcinków, posługując się RNA, jako matryca.
Kwasy rybonukleinowe
Omówimy teraz kolejny typ kwasów nukleinowych, a mianowicie kwasy rybonukleinowe.
Kwasy rybonukleinowe:
- zbudowane są z rybonukleotydów połączonych wiązaniami fosfodiestrowymi,
- występują w jądrach komórkowych i cytoplazmie,
- ich struktura zależna jest od sekwencji nukleotydów z których dana cząsteczka jest zbudowana.
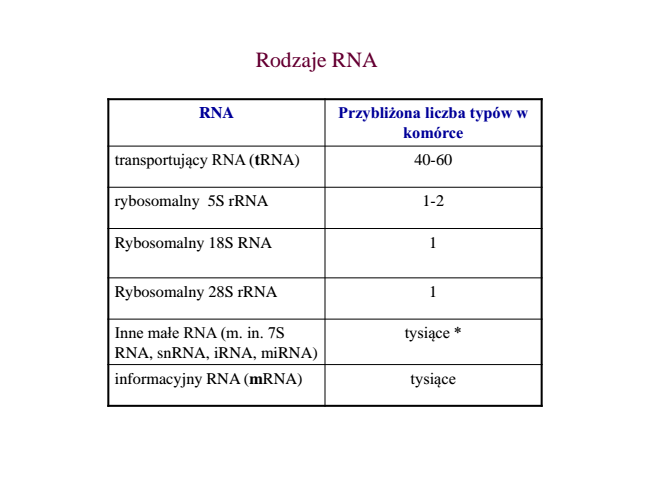
Wyróżniamy m.in.
- informacyjny/matrycowy RNA (mRNA),
- rybosomalny RNA (rRNA),
- transferowy RNA (tRNA),
- heterogenny jądrowy RNA (pre-mRNA),
- RNA regulujący ekspresję genów (interferencyjny, np. siRNA, miRNA, itd.)
RNA jest jednoniciowy, ale tworzy fragmenty dwuniciowe w obrębie siebie samego.
1) mRNA (messanger RNA)
funkcja: przenoszenie informacji genetycznej o sekwencji poszczególnych polipeptydów z genów do aparatu translacyjnego.


Powyżej przedstawiono etapy jakie musi przejść pre-mRNA. Przede wszystkim musi poddany zostać splicingowi, by usunąć z niego sekwencje niekodujące (introny). Splicing oznacza "składanie genu". Introny są bowiem z cząsteczki usuwane, a eksony łączone ze sobą. Dopiero wówczas powstanie taki mRNA, który w jednej linii będzie mógł zostać odczytany, tzn. od kodonu startu (AUG) do kodonu stopu (UAA, UAG, UAA). Proces ten jest zbędny u Procaryota, ponieważ ich geny są ciągłe w części strukturalnej, ale o tym więcej w kolejnym poście o genach i genomach.
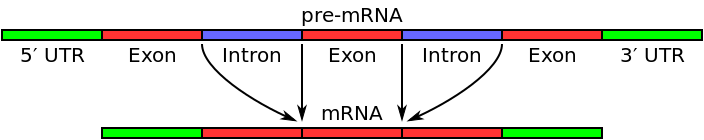
Splicing katalizowany jest kompleks białek i RNA, zwany spliceosomem, jednakże tak jak wspominaliśmy już wcześniej RNA jest zdolne do autokatalizy i może samodzielnie wycinać introny. Wówczas pełni funkcję rybozymu.
WYCINANIE PRZEZ SPLICEOSOM: atak grupy 2'OH nukleotydu adeninowego na pierwszy nukleotyd intronu na końcu 5', co powoduje powstanie pętli. Grupa 3'OH uwolnionego eksonu atakuje ostatni nukleotyd intronu na końcu 3', dzięki czemu eksony łączą się, a intron uwalniany jest w formie lassa.
SAMOWYCINANIE SIĘ INTRONÓW: atak guanozyny z intronu na miejsce splicingowe 5'. Grupa 3'OH eksonu na końcu 5' atakuje miejsce splicingowe na końcu 3', i eksony się łączą.
Łączenie eksonów niekoniecznie po kolei (losowo) nazywane jest splicingiem alternatywnym. Splicing alternatywny jest źródłem zmienności rekombinacyjnej (zmienność określana mianem mieszania genów - uzyskiwania ich nowych układów).
Na poniższym obrazku przedstawione zostały nukleotydy DNA i RNA, dla porównania, na koniec. Dodatkowo przedstawiony został schemat obróbki posttranskrypcyjnej - przyłączenia CAP'u guanozynowego i serii Adozyn - tzw łańcucha poli-A w procesie poliadenylacji końca 3'. Łańcuch Poli-A zapewnia bezpieczny transport cząsteczki mRNA z jądra do miejsca syntezy białek (do rybosomu). Z kolei CAP umożliwia przyłączenie się małej podjednostki rybosomu do mRNA. Czyli oba te "dodatki" posttranskrypcyjne umożliwiają przejście cząsteczki do kolejnego procesu - translacji - czyli tłumaczenia sekwencji nukleotydowej na sekwencję aminokwasową łańcucha polipeptydowego.

Omówiliśmy zatem strukturę mRNA, jednakże co z innymi RNA? W tym poście omówię jeszcze jeden rodzaj RNA - tRNA (transport RNA).
tRNA - RNA transportowe
tRNA to najmniejsze RNA. Jego zadaniem jest przyłączanie wolnych aminokwasów w cytoplazmie i transportowanie ich do rybosomów. Jednakże nie jest to jego jedyna funkcja. tRNA jest także czytnikiem informacji genetycznej, ponieważ zawiera tzw. antykodon na jeden z pętli, które tworzy. Antykodon decyduje o tym, jaki aminokwas transportuje. Antykodon to sekwencja trzech kolejnych nukleotydów, których zasady są komplementarne do zasad kodonu kodującego dany aminokwas na mRNA.

Więcej o tRNA pojawi się w poście dotyczącym przebiegu translacji. Omówiony zostanie wtedy cały mechanizm tworzenia łańcucha polipeptydowego. Na razie przedstawię Ci po prostu aminoacyl-tRNA - czyli tRNA "załadowane" aminokwasem. tRNA niesie te aminokwasy do rybosomu, by w mechanizmie translacji, przy udziale transferazy peptydylowej (notabene - rybozymu, ponieważ funkcję katalityczną pełni tu rRNA, czyli RNA rybosomalny dużej podjednostki rybosomu) - owe aminokwasy łączą się. Natomiast takie załadowanie aminokwasu na tRNA możliwe jest dzięki temu, że jak każdy kwas nukleinowy na końcu 3' posiada on grupę OH. Odłączamy wodór, podłączamy "aktywowany" aminokwas (dowiesz się później co to znaczy "aktywowany") i mamy - co konkretnie, jak nazwać ten twór? Aminoacylo-tRNA, w skrócie, dla leniwych - aa-tRNA.

Świetnie napisane. Pozdrawiam serdecznie.
OdpowiedzUsuńNaprawdę świetnie napisane. Pozdrawiam.
OdpowiedzUsuńEnjoy the distinctive Cascading Reels function, and have fun with the intelligent sport narrative of an alien invasion that features kidnapped cows! A good combination of Sci-Fi and slot machines will make your inner on line casino slots geek smile. Another kind of free spins bonus is when you claim them as half of} a on line casino bonus. These allow you to play slots at no cost whereas having the possibility to win real cash when you meet the connected wagering necessities. Statistically, a high-RTP online slot sport has the highest profitable odds over 온라인카지노 lengthy run|the long term}. Some of the popular options with a excessive payout share include 777 Deluxe with 96% RTP, Jungle Stripes with 97.03%, and Cyberpunk City with a powerful ninety eight.2% RTP.
OdpowiedzUsuń